Location: Na+/H+ Exchanger @ ec47b51fda2c / documentation-model.html
- Author:
- Soroush Safaei <ssaf006@aucklanduni.ac.nz>
- Date:
- 2016-05-06 15:21:46+12:00
- Desc:
- Modifying the documentation
- Permanent Source URI:
- https://staging.physiomeproject.org/workspace/28f/rawfile/ec47b51fda2c33afe8535b56355e1b9c44dedf1f/documentation-model.html
Mathematical Model
This is the general mathematical model for the Weinstein (1995) NHE3 model. See the various views available in the right hand column.
Introduction
Sodium-hydrogen antiporter 3 also known as sodium-hydrogen exchanger 3 (NHE3) or solute carrier family 9 member 3 (SLC9A3) is a protein that in humans is encoded by the SLC9A3 gene. It is found in the tubulus proximal of the nephron of the kidney and in the apical membrane of enterocytes of the intestine. It is primarily responsible for maintaining the balance of sodium. It is involved in pH regulation to eliminate acids generated by active metabolism or to counter adverse environmental conditions. NHE3 also plays an important role in signal transduction.
NHE Gene Family
The mammalian NHE (Na+/H+ exchanger) gene family (SLC9 family) consists of nine isoforms which fall into two major subdivisions: isoforms that function primarily on the plasma membrane (NHEs 1, 2, 3, 4 and 5) and those that are present primarily on intracellular organelles (NHEs 6, 7 and 9). The plasma membrane NHEs, NHE3 and 5, continually cycle between an intracellular juxtanuclear location which includes the recycling system and the plasma membrane. NHEs 1, 2 and 4 are static on the plasma membrane, although data for NHE4 are not thorough. Of the intraorganellar NHEs, NHE 6, 7 and 9 have been localized primarily to the recycling system (NHE6), trans-Golgi network (NHE7) and late endosomes (NHE9), respectively; although it is possible that these NHEs all move among intracellular organelles. The two classes of NHEs can be separated by the amino acids which make up their transport domains. NHE8 seems to resemble organellar NHEs by its amino acid composition, but it has been suggested that it resides on the plasma membrane as well. Progress has been made in understanding structure/function relationships and regulation of the plasma membrane NHEs. A major limitation to progress in the study of the organellar NHEs and NHE8 has been the failure to be able to detect organellar NHE activity in simple cell systems. Evidence from yeast studies of the endosomal (vacuolar) exchanger Nhx1 (the first organellar NHE identified) suggests that this NHE class functions as K+-Na+/H+ exchangers. These appear to act to remove intraorganellar H+ pumped in via the V-ATPase and serve to regulate organellar H+ by acting as a 'leak pathway' (Donowitz et al., 2009).
Model Structure
This transporter mediates the one-for-one exchange of Na+ for H+ and its activity is enhanced by cytosolic acidosis. Beyond mediating most of NaHCO3 reabsorption, it is also likely that the exchanger is responsible for the bulk of proximal NaCl reabsorption, functioning in parallel with Cl- anion exchange. This, along with the fact that luminal entry is rate limiting for transcellular sodium flux, renders the exchanger ideally suited as a regulatory site for proximal reabsorption. With the NHE, the primary regulatory site for proximal sodium reabsorption, flux changes will of necessity impact on cell pH, on cell volume, and on proximal ammonia transport. Thus, the exchanger is situated in a setting of sufficient complexity that a mathematical model is a natural tool for delineating the impact of transport parameters on tissue function.
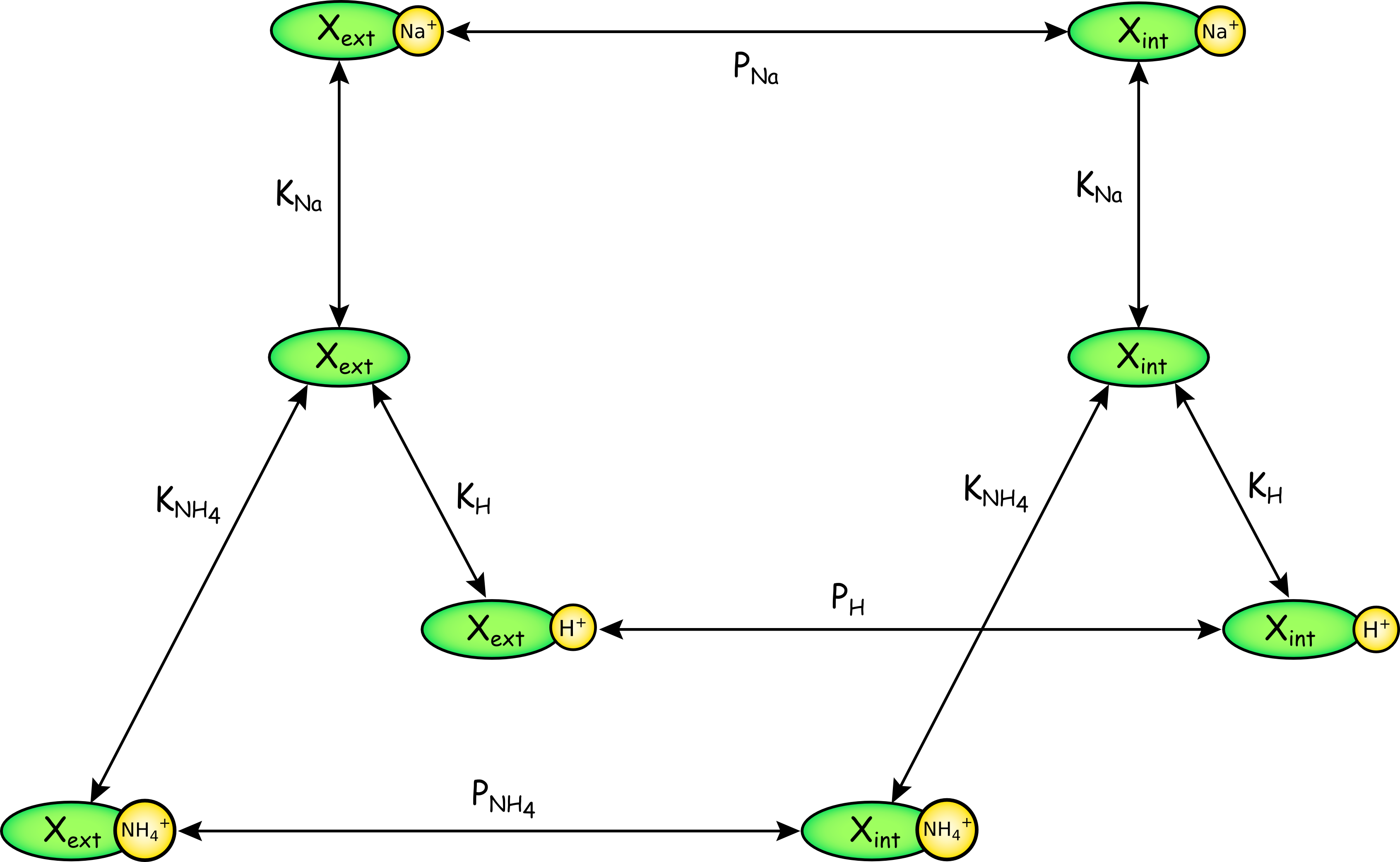
The figure above is a schematic representation of the NHE. The carrier may either be empty, X, or bound to one of three ions, Na+, H+, or NH4+. Binding is rapid, and defined by equilibrium constants KNa, KH, and KNH4. The carrier or carrier-ion complex may face either the external (ext) or internal (int) membrane surface. Translocation of the loaded carrier occurs according to rate coefficients PNa, PH, and PNH4. There is no translocation of empty carrier.
Models have heretofore relied on linear nonequilibrium thermodynamics to represent flux through the NHE. With this formulation, flux is a linear function of the transmembrane electrochemical potential differences of Na+ and H+ with a single constant transport coefficient. Once this coefficient is determined (by the overall rate of Na+ reabsorption) the kinetics are completely defined. In the present work, the approach of Heinz (1978) is used to fashion a kinetic model for the NHE. This model contains seven parameters (Na+, H+ and NH4+ affinity, the Na+, H+ and NH4+ permeation rates, and transporter density) and these are fit to the data of Aronson et al. (1983). Modification (with one additional parameter) is introduced to accommodate the observation of the internal H+ modifier site (Aronson et al., 1982). This more sophisticated transporter is then incorporated into pre-existing models of proximal epithelium and tubule (Weinstein, 1994). With these models, we illustrate the impact of the internal pH dependence for Na+/H+ exchange, on both cell volume homeostasis and overall tubule transport in metabolic alkalosis.

The figure above represents the NHE CellML model. The input variables are extracellular and intracellular concentrations of the Na+, H+ and NH4+. Permeation rates and equilibrium constants for the Na+, H+ and NH4+ are the constant parameters. The CellML model then give us the individual transmembrane solute fluxes for Na+, H+ and NH4+.
Physiological Roles of NHE3
NHE3 is an epithelial brush border NHE. NHE3 is present most abundantly in epithelial cells in the GI tract and kidney where it is responsible for the majority of Na+ absorption in these two organ systems that are responsible for most of the water and Na+ homeostasis of the body. In the intestine and proximal tubule, NHE3 functions on the apical membrane to allow absorption of large amounts of Na+ by a high capacity, low efficiency process called neutral NaCl absorption (Brett et al., 2005b).
In the intestine, Na+ absorption linked to Cl- absorption is the major way neutral NaCl absorption functions, although NHE3 activity can be uncoupled from Cl-/HCO3- exchange, at least in the mouse small intestine (Walker et al., 2008).
An important characteristic of NHE3 activity in all cell types studied is that it is in a partially activated state under basal conditions (Brett et al., 2005b). As part of normal digestive physiology, NHE3 is thought to be inhibited early in the digestive period. This presumably leads to increased luminal water and serves to spread digestive enzymes secreted by the GI organs over the intestinal surface to allow increased efficiency of digestion. Later in digestion, NHE3 activity is stimulated, to maintain volume homeostasis. In the kidney, NHE3, in addition to functioning in neutral NaCl absorption, is important for HCO3- absorption and NH4+ secretion (Bobulescu and Moe, 2006).